A nossa Terra Rara, o mais raro dos planetas no Universo, até agora único dentre todos os planetas de todos as estrelas de todas as galáxias conhecidas…
Expandindo a terapia de interferência de RNA (RNAi) do laboratório para a clínica
Otimização do projeto de siRNA para entrega do SNC no rato. (a) O siRNA direcionado a 5’Sod1 foi conjugado a lipídios através da posição 2′-O no nucleotídeo N6 da fita sense e administrado como uma única injeção de bolus de TI a ratos a 0,9’mg (com base na sequência de siRNA I). As regiões da medula espinhal e do cérebro foram coletadas 2-semanas após a dose para medição de knockdown de mRNA de Sod1 por RT-qPCR. n = 4 animais por grupo. (b) Modificação única de C16 atravessa a fita sense e a fita antisense de siRNA e transfectada em células RPE-J para avaliação de potência via RT-qPCR. n = 3 amostras biologicamente independentes. (c) C16 em várias posições de siRNA direcionado a Sod1 foi avaliado após uma única injeção de bolus de TI em ratos a 0,9 mg para knockdown na medula espinhal e regiões do cérebro por RT-qPCR. n = 3 animais por grupo. (d) Parcial ou totalmente modificado (ESC, ESC+) siRNA direcionado a Sod1 modificado com 5′-VP (Tabela Suplementar 1) foi conjugado com C16 em N6 da fita sense e administrado como uma única injeção de bolus de IT a ratos a 0,9 mg . As regiões da medula espinhal e do cérebro foram coletadas 2 semanas após a dose para medição de knockdown de mRNA de Sod1 por RT-qPCR. n = 3 animais por grupo para aCSF e parcialmente modificado; n = 5 animais por grupo para ESC; n = 2 animais por grupo para ESC+. e, o siRNA direcionado a Sod1 foi modificado com VP na extremidade 5′ da fita antisense, com 2′-O-C16 em N6 da fita sense ou com ambos e administrado como uma única injeção de bolus IT a ratos a 0,9 mg. Medula espinhal, regiões do cérebro e órgãos periféricos foram coletados 1 mês após a dose para medição de knockdown de mRNA de Sod1 por RT-qPCR. n = 3 animais por grupo. Todas as barras de erro representam o desvio padrão. (f) SiRNA direcionado a Sod1 não conjugado ou modificado por C16 foi administrado como uma única injeção de bolus IT a ratos a 0,9 mg, e a biodistribuição de siRNA foi avaliada em todo o cérebro em 24 h pós-dose usando IHC com anticorpo anti-siRNA. Três animais foram analisados por grupo com resultados semelhantes. Crédito: Nature Biotechnology (2022). DOI: 10.1038/s41587-022-01334-x
Agora é possível distribuir terapêutica baseada em RNAs interferentes curtos aos hepatócitos; no entanto, novas soluções de entrega são necessárias para atingir órgãos adicionais.
Em um novo relatório agora publicado na Nature Biotechnology, uma equipe de pesquisadores, incluindo Kirk M. Brown, Jayaprakash K. Nair e Maja M. Manas, liderada por Vasant Jadhav na Alnylam Pharmaceuticals Cambridge MA, EUA, discutiu a conjugação segura de 2′ -O-hexadecil (C16) a pequenos RNAs interferentes (siRNAs) para silenciamento potente e durável em órgãos-alvo de roedores e primatas não humanos, com ampla especificidade celular. Os experimentos forneceram atividade de interferência de RNA sustentada por pelo menos três meses. A equipe observou a administração intravítrea e intranasal, que contou com knockdown potente e durável. Eles investigaram a eficácia pré-clínica do siRNA visando a proteína precursora amilóide por meio de dosagem intracerebroventricular em um modelo de camundongo da doença de Alzheimer, que melhorou os defeitos fisiológicos e comportamentais da doença. A equipe mostrou a segurança da conjugação C16 de siRNAs para o silenciamento terapêutico de genes-alvo fora do fígado.
Terapêutica baseada em RNAi
A terapêutica de interferência de RNA (RNAi) é baseada em um mecanismo endógeno onde RNAs de interferência curtos (siRNAs) direcionam um complexo de silenciamento induzido por RNA para knockdown genético ou eliminação genética. Neste trabalho, Brown et al basearam-se em quase duas décadas de design de siRNA e otimização química para aproveitar a via de interferência de RNA em tecidos extra-hepáticos, incluindo o sistema nervoso central, olho e pulmão. Pacientes com doenças do sistema nervoso central representam algumas das maiores necessidades clínicas não atendidas com maiores desafios terapêuticos, incluindo mutações de ganho de função que os tornam adequados para silenciamento baseado em RNAi. Pesquisadores haviam realizado recentemente experimentos com siRNAs quimicamente modificados para silenciamento potente em modelos pré-clínicos por meio de uma abordagem de administração intracerebroventricular invasiva, insegura para doses repetidas em humanos.
Os bioquímicos estão atualmente desenvolvendo abordagens para permitir a entrega de siRNA através da barreira hematoencefálica que ainda estão nos estágios iniciais de descoberta. Brown et al mostraram como a conjugação de 2′-O-hexadecil (C16) pode aumentar a entrega e a captação de siRNA no epitélio alveolar e bronquiolar. Eles combinaram uma modificação lipofílica C16 com siRNAs quimicamente modificados e metabolicamente estáveis para entrega eficiente a órgãos-alvo para silenciamento de genes robusto e durável em roedores e primatas não humanos. Os experimentos forneceram um perfil de segurança favorável para gerar múltiplos candidatos para investigar a segurança e eficácia clínica.
Caracterização do C16-siRNA em SNC de rato. (a) siRNA IHC demonstrando acúmulo robusto de drogas em células neuronais e gliais (magenta) no córtex cerebral (painel esquerdo). Dual IHC para a detecção de siRNA e alvos específicos do tipo de célula em neurônios (Map2), astrócitos (Gfap) e microglia (Iba1). O sinal de siRNA em oligodendrócitos (Mbp) estava abaixo do limite de detecção de IHC. Três animais foram analisados por grupo com resultados semelhantes. (b) C16-siRNAs direcionados a transcritos específicos do tipo de célula do SNC foram administrados como uma única injeção de bolus de TI a ratos a 0,9 mg. As regiões da medula espinhal e do cérebro foram coletadas em 2 semanas após a dose (Gfap, Pecam1 e Mbp) ou 1 mês após a dose (Map2 e Iba1) para medição de knockdown de mRNA alvo por RT-qPCR. n = 3 animais por grupo para aCSF, Gfap, Iba1 e Mbp; n = 6 animais por grupo para Map2; n = 2 animais por grupo para Pecam1. (c) C16-siRNA direcionado a Sod1 foi administrado como uma única injeção de bolus de TI a ratos a 0,07 mg, 0,3 mg ou 0,9 mg. As regiões da medula espinhal e do cérebro foram coletadas em 1 mês após a dose para medição de knockdown de mRNA alvo por RT-qPCR. n = 3 animais por grupo. (d) C16-siRNA direcionado a Sod1 foi administrado como uma única injeção de bolus IT a ratos a 0,9 mg. A medula espinhal, regiões do cérebro e órgãos periféricos foram coletados nos dias indicados após a dose, até 6 meses, para medição de knockdown de mRNA alvo por RT-qPCR. n = 3 animais por grupo. (e) C16-siRNA direcionado a Sod1 foi administrado como uma injeção de bolus de IT mensal a ratos a 0,3 mg para um total de até cinco injeções ao longo de 4 meses. Medula espinhal, regiões do cérebro e órgãos periféricos foram coletados nos dias indicados, até 4,5 meses, para medição de knockdown de mRNA alvo por RT-qPCR. n = 3 animais por grupo. (f) Sod1 C16-siRNA foi administrado como injeções de TI únicas de 0,07 mg, 0,3 mg ou 0,9 mg ou injeções mensais de 0,3 mg para um total de até cinco injeções ao longo de 4 meses. As concentrações do córtex frontal mostram linearidade da dose no estudo de escalonamento de dose única em ratos e exposição aditiva no braço de múltiplas doses. A farmacocinética direta de C16-siRNA no SNC de rato é bem caracterizada por um modelo farmacocinético de primeira ordem (linhas sólidas). n = 3 animais por grupo. Todas as barras de erro representam o desvio padrão. Crédito: Nature Biotechnology (2022). DOI: 10.1038/s41587-022-01334-x
Otimizando o design de conjugados de siRNA para biodistribuição cerebral
Brown et al otimizaram cuidadosamente a lipofilicidade de siRNAs quimicamente modificados para melhorar a entrega intracelular de partículas para uma distribuição ampla e segura. Durante o processo, eles aproveitaram a posição molecular da cadeia principal de açúcar de ribose para introduzir o duplex de siRNA e monitoraram a atividade de interferência de RNA de conjugados de siRNA no sistema nervoso central. Os elementos de design de siRNA conhecidos aumentaram a potência e a especificidade no sistema nervoso central para demonstrar a melhor atividade após a combinação com vinil fosfonato e C16, com até 90 a 75 por cento de knockdown de mRNA na medula espinhal e no cérebro, respectivamente. Os dados de knockdown funcional dos conjugados mostraram biodistribuição cerebral superior após uma única injeção em ratos, em comparação com sua versão não conjugada.
Entrega eficiente de siRNA no sistema nervoso central de roedores (SNC)
Tradução da atividade de C16-siRNA em NHP CNS. (a) SiRNA direcionado a APP (compostos VII, VIII, IX e X; Tabela Suplementar 1) foi administrado como uma única injeção de bolus IT a macacos cynomolgus a 60 mg e, 3 meses após a dose, amostras de tecido foram analisadas para APP Níveis de mRNA por RT-qPCR após 85 dias. n = 5 animais por grupo para aCSF; n = 2 animais por grupo para +VP/+C16; n = 4 animais por grupo para VP/+C16; n = 1 animal por grupo para +VP/ C16; n = 3 animais por grupo para VP/C16. (b) C16-siRNA direcionado à APP foi administrado como uma única injeção de bolus de TI a três macacos cynomolgus a 60 mg. O LCR foi coletado nos dias indicados após a dose, até 9 meses, para medição de sAPP e sAPP em relação à pré-dose. n = 3 animais por grupo. (c) As concentrações de C16-siRNA foram avaliadas por espectrometria de massa de amostras de tecido do SNC obtidas de animais de aeb e de um terceiro grupo de animais (n = 3) mortos em 4,5 meses após uma dose nominal de 45 mg. Todas as barras de erro representam o desvio padrão. Crédito: Nature Biotechnology (2022). DOI: 10.1038/s41587-022-01334-x
Os bioengenheiros projetaram pequenos RNAs interferentes contra alvos específicos do tipo de célula para entender a captação de C16-siRNA e a atividade de interferência de RNA nos principais tipos de células no sistema nervoso central. Usando coloração imuno-histoquímica, eles observaram knockdown robusto em neurônios, astrócitos e células microgliais para destacar como os C16-siRNAs foram efetivamente absorvidos pela maioria dos tipos de células de relevância terapêutica no sistema nervoso central. Brown et al observaram o knockdown dose-dependente de um gene que codifica a enzima superóxido dismutase (Sod1) na medula espinhal e no cérebro e avaliaram o processo por um período de seis meses. Eles também examinaram o silenciamento dependente da dose no córtex frontal com meia-vida de siRNA por até três a quatro meses, com capacidade de examinar em um modelo farmacocinético.
O impacto potencial de pequenos RNAs interferentes (C16-siRNAs) em primatas não humanos
A equipe avaliou a potência do composto conjugado em primatas não humanos por meio de uma proteína precursora amilóide, visando o pequeno RNA interferente. O siRNA contendo vinil fosfonato e C16 mostrou melhor atividade em todo o sistema nervoso central, sem efeitos colaterais no fígado ou rim em 3 meses de terapia medicamentosa. A equipe usou amostras de líquido cefalorraquidiano para estudar a duração do silenciamento em primatas não humanos e mostrou como a variabilidade de dosagem levou à observação de uma relação robusta de resposta à exposição nas regiões do cérebro analisadas. A equipe então testou a segurança e a tolerabilidade da dose do medicamento em primatas não humanos por meio de avaliações histopatológicas de regiões do sistema nervoso central em diferentes momentos, após a administração do medicamento.
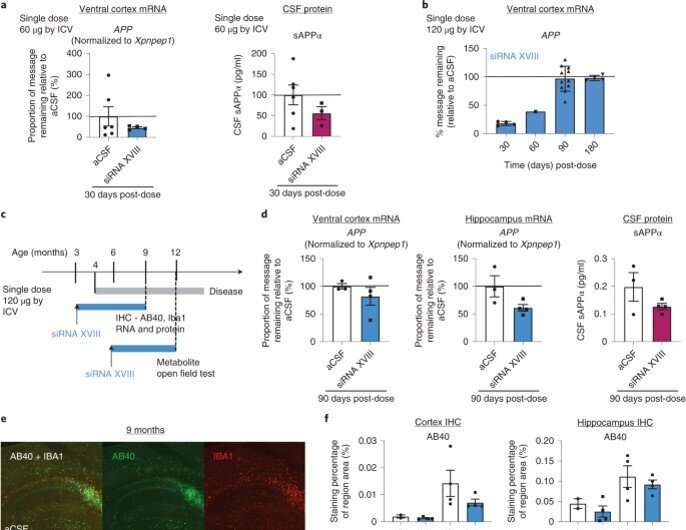
Eficácia do silenciamento de APP no modelo de camundongo CVN. (a) SiRNA XVIII direcionado à APP humana (Tabela Suplementar 1) reduziu o mRNA da APP e a proteína sAPP. aCSF, n = 6 por grupo; siRNA XVIII, n = 3 por grupo. (b) A dose única de bolus de 120 µg de ICV mostrou uma redução de ∼ 75% do mRNA de APP em 30 dias e > 50% de redução em 60 dias após a dose. Dia 30 e Dia 180, n = 4 por grupo; Dia 60, n = 1 por grupo; Dia 90, n = 11 por grupo. (c) Visão geral do desenho experimental e progressão da doença nos camundongos CVN. Os animais foram dosados pré-sintomáticos e avaliados por IHC quanto a alterações na deposição de AB40 (e,f) e inflamação (IBA1) (e,g) dentro do córtex e hipocampo 3 meses ou 6 meses após a dose. (d) Após 3 meses, uma redução de ∼25% e ∼50% do mRNA da APP foi observada no córtex e no hipocampo, respectivamente, o que correspondeu a uma redução de ∼50% na proteína sAPP. aCSF, n = 3 por grupo; siRNA XVIII, n = 4 por grupo. (f) Depósitos de Tissue AB40 avaliados pela IHC. aCSF, n = 2 por grupo em 6 meses; n = 4 para os demais grupos. (g) Níveis de IBA1 tecidual avaliados por IHC e qPCR (Iba1). aCSF, n = 2 por grupo em 6 meses; n = 4 para os demais grupos. A regressão linear simples foi usada para comparar as inclinações. *P < 0,05 (P = 0,0237 no grupo siRNA XVIII). h, os níveis de glutamato e N-acetilaspartato medidos por 1H-MRS aos 12 meses de idade (6 meses pós-dose) mostram normalização dos níveis de glutamato no grupo tratado com siRNA. WT aCSF, n = 9 por grupo; n = 8 por grupo para os demais grupos. Todas as barras de erro representam o erro padrão. *P< 0,05. Foi usado o teste t não pareado assumindo variância igual. CR, creatina i, animais tratados com siRNA mostram normalização da distância total percorrida e frequência de criação. WT aCSF, n = 9 por grupo; n = 8 por grupo para os demais grupos. Todas as barras de erro representam o erro padrão. *P < 0,05, **P < 0,005. O teste t não pareado assumindo variância igual foi usado a menos que indicado de outra forma. NS, não significativo. Crédito: Nature Biotechnology (2022). DOI: 10.1038/s41587-022-01334-x
Distribuição e atividade de C16-siRNA no pulmão de camundongo. (a,b) C16-siRNAs foram administrados em camundongos por via intranasal a 10 mg kg 1 (Sod1) (a) ou 30 mg kg 1 (Traf6) (b). Os pulmões foram coletados no dia 10 ou dia 28 pós-dose, respectivamente, para siRNA IHC. siRNA, magenta; Contracoloração de hematoxilina, azul. Três animais foram analisados por grupo com resultados semelhantes. (c) O knockdown de mRNA de Sod1 foi visualizado por ISH no dia 10 após uma dose de 10 mg kg 1 IN de C16-siRNA direcionado a Sod1. mRNA de Sod1, marrom; Contracoloração de hematoxilina, azul. Três animais foram analisados por grupo com resultados semelhantes. (d) RT qPCR de mRNA de Sod1 no pulmão inteiro medido no dia 10 após 0,3, 1, 3 ou 10 mg kg 1 dose única IN ou 30 mg kg 1 dose IV única de C16-siRNA direcionado a Sod1. n = 3 animais por grupo. Todas as barras de erro representam o desvio padrão. (e) RT qPCR de mRNA de Sod1 no pulmão inteiro medido em vários pontos de tempo após a dose de 10 mg kg 1 IN de C16-siRNA direcionado a Sod1. n = 3 animais por grupo. Crédito: Nature Biotechnology (2022). DOI: 10.1038/s41587-022-01334-x
Eficácia pré-clínica do composto conjugado em um modelo de camundongo
Brown et al usaram camundongos transgênicos com mutações específicas para a doença de Alzheimer para avaliar a eficácia pré-clínica do composto desenvolvido durante a pesquisa. A equipe mostrou como uma dose única (60 µg) do composto reduziu os níveis de proteína mutante no córtex ventral e no líquido cefalorraquidiano, em um período de tempo variável. Os cientistas estudaram a dinâmica da administração do composto e usaram a espectroscopia magnética de prótons para determinar a diminuição da deposição de proteínas e dos níveis de inflamação, para restaurar a função neuronal. A equipe observou a redução de metabólitos específicos em uma cepa de camundongos CVN, onde o knockdown de genes específicos normalizou metabólitos específicos para trazê-lo de volta ao mesmo nível da cepa do tipo selvagem. Os pesquisadores demonstraram ainda a atividade potente e durável do composto no olho e no pulmão.
Perspectivas: evolução do tratamento clínico
Desta forma, Kirk M. Brown, Jayaprakash K. Nair e Maja M. Manas e colegas projetaram pequenas interferências (siRNAs) para aplicações extra-hepáticas. A equipe combinou conjugados de siRNA lipofílicos com recursos de design adicionais para ampla distribuição no sistema nervoso central de modelos animais pré-clínicos por meio de uma rota de dosagem segura clinicamente favorável. Eles conduziram um estudo de prova de conceito em um modelo de camundongo pré-clínico para normalizar déficits comportamentais, com resultados adequados para desenvolvimentos clínicos para tratar a doença de Alzheimer de início precoce e a angiopatia amilóide cerebral. A estratégia de conjugação também é aplicável no olho e no pulmão após a entrega local. A equipe realizará mais estudos para entender o mecanismo preciso por trás da entrega intracelular de C16 para absorção em vários tipos de células.