A nossa Terra Rara, o mais raro dos planetas no Universo, até agora único dentre todos os planetas de todos as estrelas de todas as galáxias conhecidas…
Cientistas ‘acidentalmente’ criaram embriões de frango com rostos de dinossauros
Se os filmes de “Jurassic Park” nos ensinaram alguma coisa, é que definitivamente não devemos tentar ressuscitar dinossauros. Será??
Embora pareça impossível trazê-los de volta, já que quase todas as suas espécies foram exterminadas há cerca de 65 milhões de anos, os cientistas apenas provaram que algumas de suas características físicas podem realmente ser replicadas nos descendentes modernos de dinossauros emplumados, o único grupo que sobreviveu à extinção . Mas não é preciso ter medo de tiranossauros rexes e velociraptores correndo desenfreados porque as únicas coisas com as quais você precisa se preocupar são galinhas com rostos de dinossauros. Sim, você leu corretamente.
Sessenta e cinco milhões de anos atrás, acredita-se que um asteróide tenha colidido com a Terra. O impacto eliminou um grande número de espécies, incluindo quase todos os dinossauros.
Velociraptores eram pequenos dinossauros terópodes de penas (Crédito: Sabena Blackbird / Alamy)
Um grupo de dinossauros conseguiu sobreviver ao desastre. Hoje, nós os conhecemos como pássaros.
Para entender como um se transformou em outro, uma equipe vem mexendo nos processos moleculares que compõem um bico em galinhas.
Ao fazer isso, eles conseguiram criar um embrião de galinha com um focinho e palato parecido com um dinossauro, semelhante ao de pequenos dinossauros de penas como o Velociraptor. Os resultados são publicados na revista Evolution.
O objetivo da equipe era entender como o bico do pássaro evoluiu, porque o bico é uma parte vital da anatomia do pássaro. Foi crucial para o sucesso deles. As 10.000 ou mais espécies de aves ocupam uma grande variedade de habitats e muitas possuem bicos especializados para ajudá-las a sobreviver.
Mas eles não se propuseram a criar uma “galinha dino”, dizem os principais autores Bhart-Anjan Bhullar, da Universidade de Yale, em New Haven, e Arkhat Abzhanov, da Universidade de Harvard, em Cambridge, EUA.
O primeiro fóssil de Archaeopteryx foi descoberto em 1861 (Crédito: Age fotostock / Alamy)
“Sempre que você examina uma importante transformação evolutiva, deseja aprender o mecanismo subjacente”, diz Bhullar.
O bico também é a parte do esqueleto aviário que “se diversificou mais extensivamente e mais radicalmente”, diz Bhullar.
Apesar dessa diversidade – variando de flamingos a pelicanos – muito pouco trabalho foi feito para descobrir “o que diabos é um bico”, acrescenta.
“Eu queria saber o que o bico era esquelético, funcional e quando essa grande transformação ocorreu de um focinho de vertebrado normal para as estruturas únicas usadas nas aves”.
Galinhas são parentes distantes de dinossauros (FL Collection / Alamy)
Para começar a entender isso, a equipe vasculhou as mudanças na maneira como os genes são expressos nos embriões de galinhas e em vários outros animais. Eles examinaram os embriões de ratos, emas, jacarés, lagartos e tartarugas, representando muitos dos principais grupos de animais.
Eles descobriram que os pássaros têm um conjunto único de genes relacionados ao desenvolvimento facial, que as criaturas não-bicudas não possuíam.
Quando eles silenciaram esses genes, a estrutura do bico voltou ao seu estado ancestral. O mesmo aconteceu com o osso palatino no céu da boca.
Controle de embriões de galinha, embrião de galinha alterado e embrião de jacaré
Para fazer esse ajuste genético, Bhullar e seus colegas isolaram as proteínas que teriam desenvolvido os bicos. Em seguida, os suprimiram usando pequenas contas revestidas com uma substância inibidora.
Quando seus esqueletos começaram a se desenvolver dentro dos ovos, esses animais tinham ossos curtos e arredondados em vez de bicos alongados e fundidos que os esqueletos de pássaros.
“Ao afetar essa proteína inicial, você está realmente alterando a expressão do gene”, acrescentou Bhullar.
O trabalho destaca que os bicos se desenvolvem de maneira muito diferente dos focinhos, usando um conjunto diferente de genes, diz Michael Benton, da Universidade de Bristol, no Reino Unido. “Isso é o que prova que o bico é uma adaptação ou ‘coisa’ real, não apenas uma forma ligeiramente diferente do nariz”.
A mudança dos focinhos para os bicos aconteceu bem na evolução das aves, 40-50 milhões de anos após o Archaeopteryx, diz Benton.
Os bicos de pássaros têm várias formas e tamanhos (Crédito: John Warburton-Lee / Alamy)
Por enquanto, Bhullar não tem planos ou aprovação ética para chocar as galinhas de focinho. Mas ele acredita que eles poderiam sobreviver “muito bem”.
“Essas não foram modificações drásticas”, diz Bhullar. “Eles são muito menos estranhos do que muitas raças de frango desenvolvidas por amadores e criadores de frango”.
“O resto do animal parecia bem, mas é preciso pensar com cuidado sobre isso do ponto de vista ético.”
Como foi o experimento da “Galinhassaura”:
Usando uma substância inibidora, eles conseguiram suprimir as proteínas que originariam o bico. O resultado é que obtiveram um embrião de frango com cara de dinossauro, ou seja, com aquele “focinho” e sem o bico. Quando silenciaram esses genes, a estrutura do bico voltou ao seu estado ancestral. Assim também o osso palatino no teto da boca.
O “experimento” criou um embrião de frango semelhante aos pequenos dinossauros emplumados como Velociraptor. Apesar de não terem dentes, os embriões apresentavam um focinho largo, arredondado e com irregularidades. O objetivo não era conseguir uma galinha bizarra e os autores do estudo estão chamando a descoberta de “acidental”.
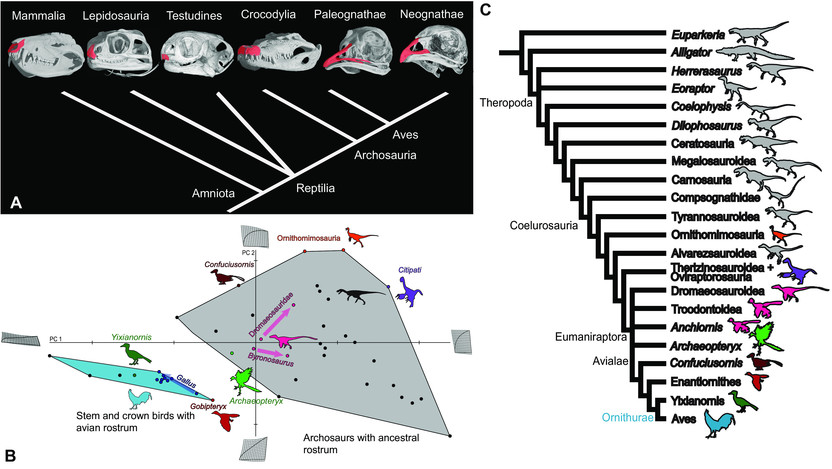
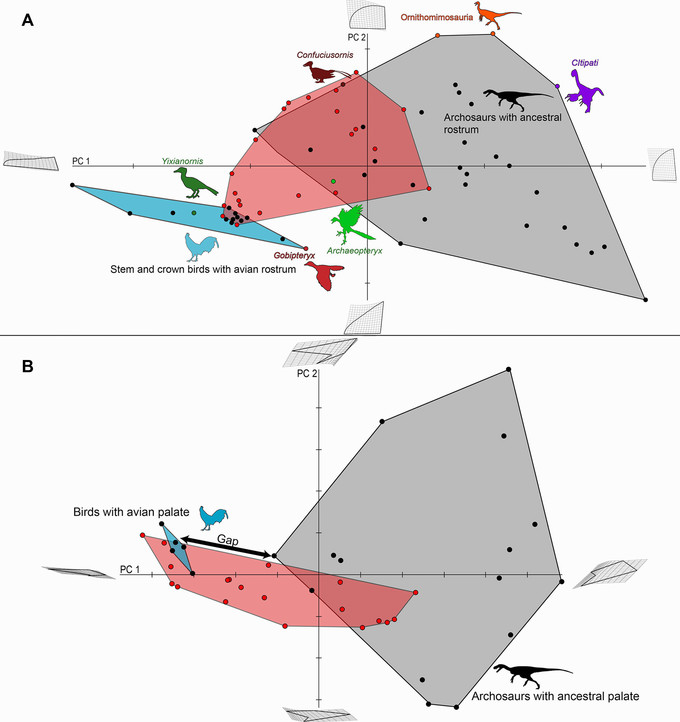
A origem da tribuna aviária. (A) Filogenia amniote mostrando principais clados e configuração das pré-maxilas. O rostro aviário é formado a partir de pré-maxilas alongadas, autapomorficamente alongadas. Imagens de UTCT / Digimorph.org. (B) Gráfico de PCA da vista dorsal da pré-maxila com imagens de extremos hipotéticos ao longo de cada eixo, dispostas em grades de deformação a partir da média. Os táxons com tribuna aviária, incluindo a forma transitória Yixianornis e galinhas adultas e embrionárias (em azul escuro), aglomeram-se longe dos arquossauros restantes. Gobipteryx evoluiu de forma convergente pré-maxila aviária. Confuciusornis, ornitomimosauros e alguns oviraptorossauros (Citipati aqui) não tinham dentes e provavelmente tinham uma rhamphotheca, ambos componentes do “bico” definitivo das aves, mas a forma pré-maxilar não era parecida com um pássaro. Trajetórias ontogenéticas aproximadas são mostradas usando setas para dromaeossauros em geral, Byronosaurus e Gallus. Os dois dinossauros não-aviários têm premaxilas mais longas e mais estreitas quando mais jovens; o pássaro tem premaxilas mais longas e mais estreitas quando mais velhas. (C) Filogenia de Archosauria mais Euparkeria mostrando os principais clados discutidos no texto. Modificado após Bhullar et al. (2012).
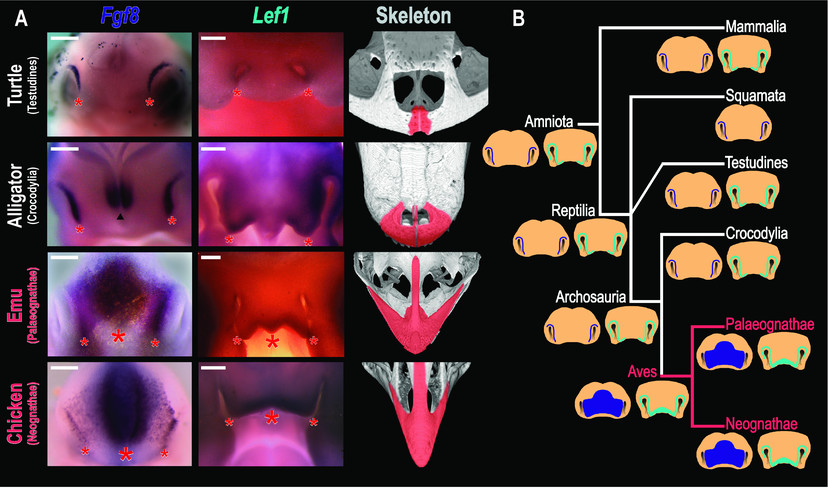
Expressão de genes de padrões faciais em amniotes. (A) Expressão de Fgf8 e Lef1 e fenótipos esqueléticos adultos em répteis, mostrando expressão ancestral de genes emparelhados precedendo pequenas pré-maxilas emparelhadas em tartarugas e jacarés e zona mediana de expressão anterior a premaxilas alongadas e fundidas em aves. Imagens esqueléticas de UTCT / digimorph.org. (B) Filogenia amniote com dados sobre a expressão do gene de padrão facial e estados ancestrais inferidos mostrados. Azul escuro: Fgf8. Azul claro: Lef1. A zona de expressão mediana é uma autapomorfia das aves correlacionada com a presença do rostro aviário. Barras de escala 500 ?m.
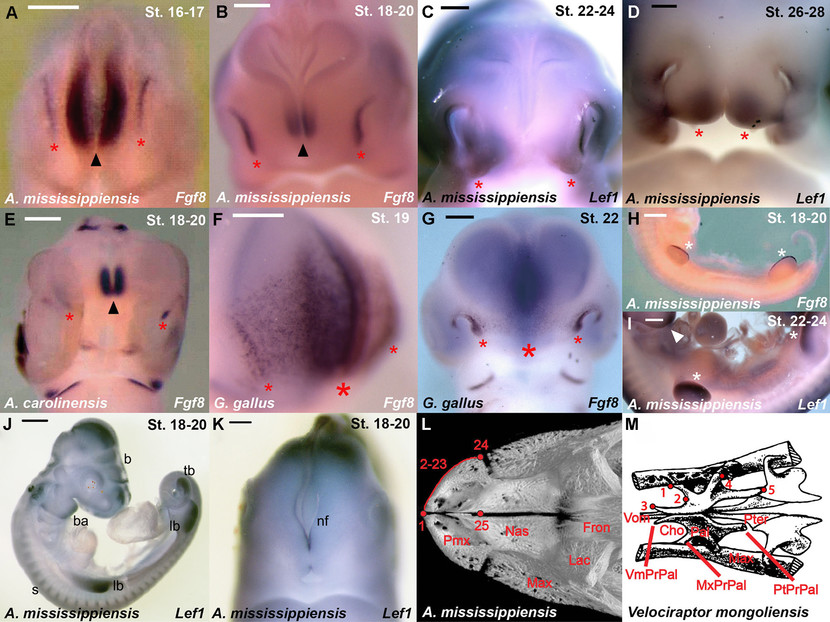
Dados adicionais de expressão gênica e posições de referência. (A) Expressão de Fgf8 (estrelas vermelhas) em jacaré equivalente a HH 16-17, mostrando expressão epitelial ancestral de amniote ancestral, mesmo em estágio inicial; ponta de seta indica expressão no cérebro. (B) Expressão de Fgf8 (estrelas vermelhas) em jacaré equivalente a HH 18-20, mostrando expressão emparelhada com ponta de seta indicando expressão no cérebro. (C) Expressão de Lef1 em jacaré equivalente a HH 22–24, mostrando expressão emparelhada de amniote ancestral sem expressão na linha média. (D) Expressão de Lef1 no jacaré equivalente a HH 26–28, mostrando ausência contínua de expressão da linha média na região onde as aves apresentam crescimento decrescente do ápice do bico presuntivo (consulte A). Eventualmente, a sutura entre as pré-maxilas será formada aqui. (E) Expressão de Fgf8 no lagarto anole equivalente a HH 18-20, mostrando expressão pareada de amniote ancestral (estrelas) com ponta de seta indicando a expressão do cérebro. (F) Expressão de Fgf8 em frangos HH 19, mostrando ampla faixa mediana de expressão exclusiva para aves. (G) Expressão de Fgf8 em galinha HH 22, mostrando expressão contínua na linha média em estágio relativamente tardio. (H) expressão de Fgf8 no corpo do jacaré equivalente a HH 18–20 (lateral esquerdo, invertido) mostrando expressão nas cristas ectodérmicas apicais (estrelas). (I) Padrão de expressão de Lef1 no corpo do jacaré equivalente a HH 22–24 (lateral direito) mostrando acúmulo de mRNA nos membros (estrelas), somitos e arcos branquiais (ponta da seta). (J) Expressão de Lef1 em jacaré equivalente a HH 18-20, mostrando atividade distinta nas regiões esperadas do cérebro (b), arcos branquiais (ba), gemas de membros (lb), gema de cauda (tb) e somitos (s) . (K) Detalhe da cabeça do jacaré equivalente a HH 18–20 corado para expressão de Lef1. Não há expressão discernível no mesênquima facial anterior; assim, a atividade WNT segue temporalmente a atividade FGF na frente da face. A expressão é claramente visível nas dobras neurais (nf) do cérebro anterior. (L) Marcos pré-maxilares e anatomia facial na tribuna de jacaré em vista dorsal. Pmx, pré-maxila; Nas, nasal; Max, maxila; Lac, lacrimal. (M) Marcos palatinos e anatomia palatal no Velociraptor mongoliensis modificados após (Barsbold e Osmólska 1999). Vom: Vomer. Pal: Palatino. Pterigoide. Max: Maxilla. Cho: margem choanal. VmPrPal: processo vomerino de palatino. MxPrPal: Processo maxilar do palato. PtPrPal: Processo pterigóide de palatina. Barras de escala de 500 ?m (a-g, j) e 1 mm (h-i, k).
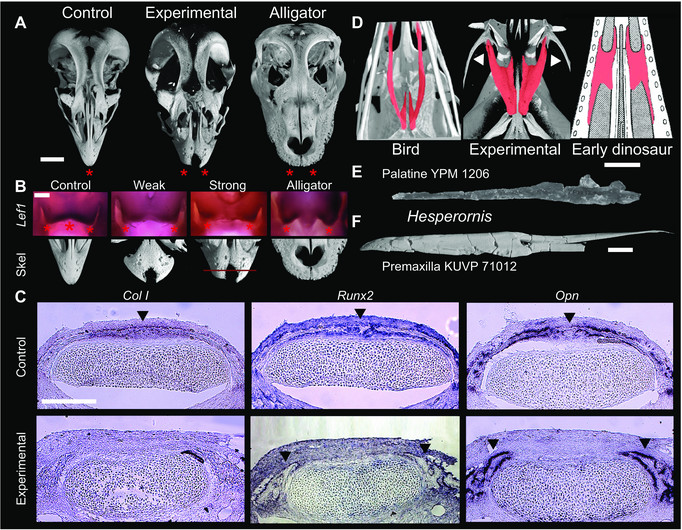
Restauração experimental da expressão ancestral prevista. (A) Frango controle, frango experimental e jacaré, mostrando pré-maxilares ancestrais emparelhadas, abreviadas e arredondadas no animal experimental. (B) Expressão de Lef1 (estrelas vermelhas) em frangos de controle, experimentais e jacarés e fenótipos pré-maxilares correspondentes. Nos animais experimentais inibidos por Fgf8, a expressão de Lef1 é mais pareada como o estado ancestral representado pelo jacaré. (C) Expressão de marcadores esqueléticos e esqueletogênicos em embriões controle e experimental; nível de seção mostrado em vermelho na imagem do esqueleto do fenótipo forte. A zona mediana de fusão que une as duas pré-maxilas nas galinhas de controle (pontas superiores da seta) está ausente nos experimentais, que em vez disso têm ossificações emparelhadas (pontas inferiores da seta). (D) Morfologia palatal em galinha normal (UTCT / digimorph.org), embrião experimental e arquossauro ancestral (após Colbert 1989). O palato no embrião experimental se assemelha à forma ancestral por ter um processo maxilar curto terminando na maxila (pontas de seta) e uma lâmina triangular extensa entre os processos. (E) O palato do pássaro-tronco Hesperornis regalis YPM (Museu Yale Peabody) 1206. Este elemento é essencialmente idêntico ao dos pássaros do tipo coroa em sua configuração geral. (F) As pré-maxilas fundidas da ave caule Hesperornis regalis KUVP (Universidade do Kansas) 71012. Como o palatino, esse elemento representa essencialmente uma configuração aviária moderna. Barras de escala de 1 cm (a, e, f), 500 ?m (b), 100 ?m (c).
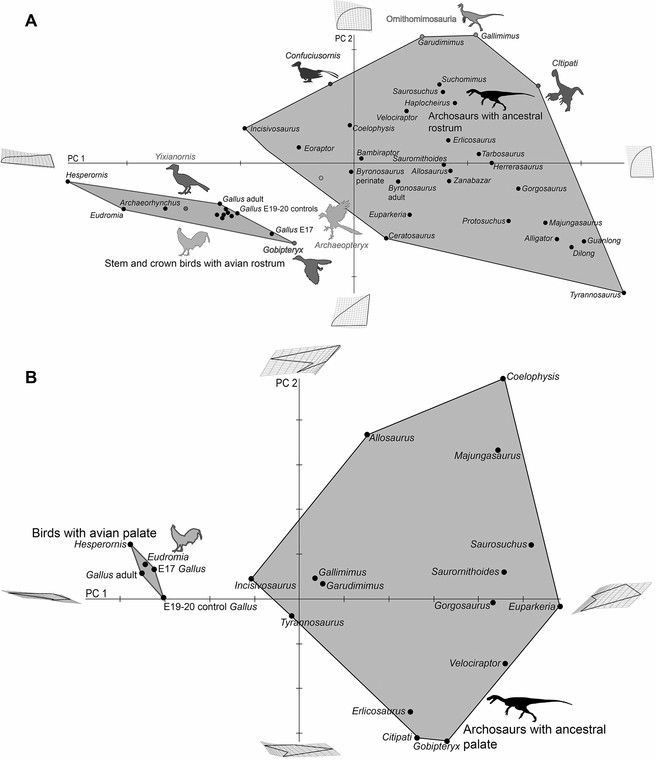
Análise morfométrica de embriões experimentais. (A) PCA de pré-maxilas experimentais, mostrando que os experimentais (em vermelho) se sobrepõem parcialmente ao morfoespaço arquossônico ancestral. (B) PCA de palatinos experimentais mostrando que os experimentais (em vermelho) preenchem a lacuna entre as aves do grupo da coroa e os arquossauros mais ancestrais e se sobrepõem ao morfoespaço dos araras ancestrais não aviais. Prevemos que, à medida que os primeiros palatinos avialanos se tornarem mais conhecidos, os fósseis ocuparão o espaço entre os fenótipos experimentais.
Assine nossa newsletter e fique informado sobre Astrofísica, Biofísica, Geofísica e outras áreas. Preencha seu e-mail no espaço abaixo e clique em “OK”: