A nossa Terra Rara, o mais raro dos planetas no Universo, até agora único dentre todos os planetas de todos as estrelas de todas as galáxias conhecidas…
Engenharia de ácidos ribonucleicos circulares (circRNAs) para melhorar a produção de proteínas
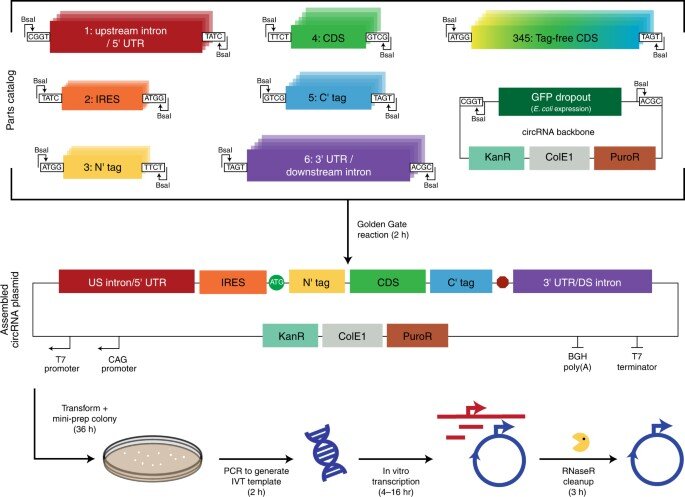
Uma plataforma de clonagem modular para circRNA permite ciclos rápidos de design-construção-teste. Esquema que descreve a plataforma de clonagem modular usada para criar plasmídeos modelo para a síntese de circRNA. As partes 1-6 correspondentes ao íntron a montante e 5′ UTR, IRES, tag N-terminal (N’), sequência de codificação (CDS), tag C-terminal (C’) e 3′ UTR e o íntron a jusante foram clonados individualmente em parte plasmídeos através de reações Golden Gate (Fig. 1 suplementar). Os plasmídeos de parte e o backbone circRNA foram então combinados em uma segunda reação Golden Gate para criar um plasmídeo circRNA. A estrutura de circRNA contém um promotor CAG que permite a transcrição de circRNA após a transfecção transitória em cellulo, um promotor T7 que permite IVT, sequências de homologia que auxiliam na circularização de RNA, regiões de baixa estrutura que facilitam a processabilidade de RNaseR e uma sequência de eliminação de GFP expressa por bactérias para selecionar negativamente montagens. Se um CDS sem tags N’ ou C’ foi usado, as partes 3-5 foram substituídas por uma única parte. Os produtos de PCR de plasmídeos circRNA foram subsequentemente usados como moldes para IVT para sintetizar RNA. Por fim, a limpeza de RNaseR foi realizada para digerir RNAs lineares e isolar circRNA. DS, a jusante. Crédito: Nature Biotechnology (2022). DOI: 10.1038/s41587-022-01393-0
Os ácidos ribonucleicos circulares (circRNAs) são uma plataforma promissora para estudos de expressão gênica como um ácido ribonucleico estável e prevalente em células eucarióticas, que surgem de back-splicing. Em um novo relatório agora publicado na Nature Biotechnology, Robert Chen e uma equipe de pesquisadores interdisciplinares da Universidade de Stanford, Califórnia, EUA, desenvolveram uma abordagem sistemática para montar e testar rapidamente recursos que afetam a produção de proteínas com base em RNAs circulares sintéticos.
A equipe maximizou a tradução do circRNA otimizando elementos finos para implementar princípios de design para melhorar o rendimento de RNA circular em várias centenas de vezes. Os resultados facilitaram um aumento da tradução do RNA de interesse, quando comparado aos níveis de RNA mensageiro (mRNA), para fornecer tradução durável in vivo.
Desenvolvimento de RNA circular (circRNA) no laboratório
A terapêutica baseada em ácidos ribonucleicos abrange RNA mensageiro (mRNA), pequenos RNAs interferentes (siRNA) e microRNAs (miRNA) com expansão para a medicina moderna, incluindo pequenas moléculas, produtos biológicos e terapia celular. Por exemplo, as vacinas de mRNA recentemente populares podem ser projetadas em laboratório e desenvolvidas em ritmo acelerado para responder a crises médicas urgentes e em evolução. Os RNAs de codificação podem ser circularizados em circRNAs para estender a duração da tradução de proteínas, com base em moléculas de RNA que se unem covalentemente cabeça-cauda. Os bioengenheiros também avançaram na síntese de transcritos longos circulares em circRNAs. No entanto, os mecanismos fundamentais de iniciar a tradução para formar RNA circular ou RNA mensageiro diferem devido à falta de uma capa de 7-metilguanilato (M7G) nos RNAs circulares. Como resultado disso, os pesquisadores precisam examinar minuciosamente os princípios da tradução circular de RNA para construir melhores terapias e potencialmente superar as capacidades de tradução do mRNA. Para examinar esse aspecto, a equipe desenvolveu uma plataforma modular de alto rendimento para construir e testar RNAs circulares sintéticos para tradução otimizada e rendimentos de proteína aprimorados.
A otimização de elementos não codificantes de RNA permite uma tradução mais forte de circRNA. (a) Atividade NanoLuc após transfecção de células HeLa com circRNAs contendo um IRES 3′ ou 5′ e sequências espaçadoras de comprimentos variados. Quando o IRES está a 3′ do repórter NanoLuc, a tradução através da cicatriz td splicing é inevitável. A estrutura secundária prevista desta cicatriz é mostrada. A atividade do NanoLuc foi normalizada para a atividade constitutiva da luciferase do vaga-lume da mesma amostra e depois dividida pelos valores da transfecção simulada. Os dados são média ± s.e.m. para n = 3 réplicas biológicas. (b) Atividade NanoLuc em 24 horas após a transfecção de células HeLa com circRNAs contendo o número indicado de códons de parada. A atividade do NanoLuc foi normalizada para a atividade constitutiva da luciferase do vaga-lume da mesma amostra e depois dividida pelos valores da transfecção simulada. Os dados são média ± s.e.m. para n = 4 réplicas biológicas. (c) Atividade de NanoLuc após transfecção de células HeLa com circRNAs contendo diferentes sequências espaçadoras 5′. A atividade do NanoLuc foi normalizada para a atividade constitutiva da luciferase do vaga-lume da mesma amostra e depois dividida pelos valores da transfecção simulada. Os dados são média ± s.e.m. para n = 3 réplicas biológicas. *P = 0,0213, **P = 0,0051 e ***P < 0,001 por teste t bilateral não pareado em comparação com uma sequência espaçadora aleatória de 50 nt. (d) Atividade NanoLuc após transfecção de células HeLa com circRNAs contendo diferentes sequências 3' UTR. A atividade do NanoLuc foi normalizada para a atividade constitutiva da luciferase do vaga-lume da mesma amostra e depois dividida pelos valores da transfecção simulada. Os dados são média ± s.e.m. para n = 3 réplicas biológicas. ***P = 0,0012 e ****P < 0,0001 pelo teste t bilateral não pareado em comparação com uma sequência espaçadora aleatória de 50 nt. BR, região de ligação; RM, região mínima; PR, região protegida. Crédito: Nature Biotechnology (2022). DOI: 10.1038/s41587-022-01393-0
Uma plataforma modular de montagem circRNA
Os cientistas desenvolveram uma plataforma de clonagem modular feita de um conjunto de peças compatível com a clonagem Golden Gate e Gibson para permitir testes de maior rendimento de circRNAs. Usando a plataforma, eles determinaram como aspectos específicos do design de RNA circular afetaram sua tradução. Por exemplo, a equipe havia mostrado anteriormente como o RNA circular desencadeado respostas imunes pode ser evitado in vivo, modificando as moléculas com m6A. No entanto, os pesquisadores ainda precisam entender o impacto dessa etapa na tradução de RNA circular. Para resolver isso, Chen e a equipe usaram sua plataforma de clonagem e incorporaram o m6A. Quando comparados com circRNAs não modificados, aqueles contendo 5% de m6A mostraram tradução igual após transfecção ou eletroporação in vitro. Os cientistas, assim, avaliaram experimentalmente o impacto da modificação na estabilidade do RNA circular.
CircRNAs projetados demonstram tradução mais durável e atividade funcional in vivo. (a) CircRNA com 5% de incorporação de m6A que codifica NanoLuc foi sintetizado com as seguintes otimizações: topologia IRES a montante, espaçador PABP 5′, HBA1 3′ UTR e HRV-B3 IRES com inserção de loop proximal Apt-eIF4G. CircRNAs foram formulados para entrega intraperitoneal em camundongos usando CARTs. A expressão foi testada usando um sistema de imagem óptica após injeções intraperitoneais do substrato de fluorofurimazine nos pontos de tempo indicados. Em 336 horas (14 dias) após a administração do circRNA NanoLuc, os camundongos foram redosados. (b) Imagem luminescente in vivo de um camundongo não tratado (esquerda) versus camundongos recebendo circRNA NanoLuc (direita) em 24 horas após a dosagem. (c) Quantificação de luminescência por camundongo em diferentes momentos após a administração de circRNA NanoLuc. A redosagem foi realizada em 336 horas (14 dias). Os dados são média ± s.e.m. para n = 3 animais por condição. (d) CircRNA com 5% de incorporação de m6A que codifica hEPO foi sintetizado com as seguintes otimizações: topologia IRES a montante, espaçador PABP 5′, HBA1 3′ UTR e HRV-B3 IRES com inserção de loop proximal Apt-eIF4G. A hEPO codificadora de mRNA foi sintetizada com reagente CleanCap, 100% de incorporação de N1 e uma cauda poli(A) de 120 nt. Doses equimolares de circRNA e mRNA foram formuladas para entrega intravenosa em camundongos usando CARTs. Plasma hEPO foi medido por ELISA em uma coorte nos pontos de tempo indicados. Os reticulócitos foram contados em uma coorte separada em 168 horas (7 dias). (e) Quantificação de hEPO no plasma em diferentes momentos após a administração de circRNA hEPO ou mRNA hEPO. Os dados são média ± s.e.m. para n = 4 animais por condição. (f) Expressão de hEPO no plasma normalizada para o nível de 24 horas de cada camundongo em diferentes pontos de tempo após a administração de circRNA hEPO ou mRNA hEPO. Os dados são média ± s.e.m. para n = 4 animais por condição. *P = 0,0487 e ***P = 0,0001 pelo teste t bilateral não pareado com correção de Bonferroni comparado ao mRNA. g, porcentagem de reticulócitos entre os glóbulos vermelhos em 168 horas após a administração de circRNA hEPO ou mRNA hEPO. Os dados são média ± s.e.m. para n = 4 animais por condição. **P = 0,0080 pelo teste t bilateral não pareado. NS, não significativo. Crédito: Nature Biotechnology (2022). DOI: 10.1038/s41587-022-01393-0
CircRNA exibe tradução mais durável do que mRNA in vitro. Atividade de NanoLuc no sobrenadante após eletroporação de células HeLa com circRNA ou mRNA que codifica NanoLuc secretado. CircRNA foi sintetizado com 5% de incorporação de m6A e o HRV-B3 IRES. O mRNA foi sintetizado com reagente CleanCap, 100% de incorporação de N1 e uma cauda poli(A) de 120 nt. Nas horas indicadas (h) e dias (d) pós-eletroporação, o meio foi colhido para testar o NanoLuc secretado e substituído. A atividade do NanoLuc foi dividida pelos valores da eletroporação simulada. Os dados são média ± SEM para n=3 réplicas biológicas. Crédito: Nature Biotechnology (2022). DOI: 10.1038/s41587-022-01393-0
Descobrindo a dinâmica do circRNA para resultados de tradução fortes
Para descobrir os princípios subjacentes à topologia do vetor circRNA necessária para uma tradução forte, os pesquisadores começaram a sintetizar circRNAs para gerar variantes com peptídeos codificados pelo processo. Com base nos resultados, a equipe mostrou que aumentar o comprimento do espaçador não era benéfico para a tradução. Em seguida, eles mostraram como as regiões não traduzidas 5′ e 3′ podem melhorar a tradução do circRNA. Os pesquisadores também conduziram uma série de experimentos para examinar a otimização do circRNA e depois os compararam em um único experimento. Eles mostraram como as mudanças aumentaram progressivamente a expressão do circRNA sem comprometer o rendimento do RNA ou a eficiência da circularização. Eles também mostraram como a cinética da tradução do circRNA e do mRNA difere significativamente, onde o circRNA levou mais de 24 horas para atingir seu comprimento máximo de tradução, excedendo em muito a duração da tradução do mRNA. Eles então combinaram a série de otimizações de circRNA para testar sua expressão in vivo. Para entregar os RNAs, a equipe os formulou com transportadores liberáveis de alteração de carga (CARTs) ou moléculas catiônicas que mediam a expressão de mRNA em modelos de camundongos. Os resultados mostraram como os circRNAs modificados podem ser expressos em forças semelhantes aos RNAs modificados in vivo, embora com maior duração.
Panorama
Dessa forma, Robert Chen e colegas mostraram como a circularização de RNA tem grande potencial para transformar medicamentos baseados em RNA, estendendo a durabilidade de moléculas relativamente altamente transitórias. Dadas as diferenças fundamentais entre os mecanismos de tradução de circRNA e mRNA, o conhecimento existente de maximizar a tradução de mRNA não necessariamente traduziu para circRNAs. Para facilitar este estudo, a equipe criou uma plataforma de clonagem modular circRNA para testar inúmeras variações de sequência e otimizações de vários parâmetros. Usando a plataforma, eles identificaram várias abordagens para melhorar a tradução de proteínas de circRNAs, com aplicações para engenharia ampla de RNAs, para produzir mais proteínas do que mRNAs in vitro e exibir maior durabilidade de sua tradução in vivo e in vitro. Os cientistas dissecaram sistematicamente os elementos que regulam a tradução do circRNA para otimizar as regiões de interesse para aumentar o rendimento da proteína circRNA para a produção de proteínas duráveis in vivo.